1-12 of 22 results
-

How to respond to the question “Should I take a probiotic?”
Two experts set the record straight on a media article about the benefits of probiotics. -

Discovering novel bioactive peptides in fermented foods
Food not only serves as a primary source of essential nutrients but also contains a wealth of potential bioactive compounds…. -

A guide to the new FDA Qualified Health Claim for yogurt
Fermented foods such as yogurt, kimchi, and fermented pickles have traditionally been associated with health benefits in countries around the… -

Fermented Food Microbiology Researcher in Mohali, India Receives 2024 Gregor Reid Award for Outstanding Scholars in Developing Nations
ISAPP’s board of directors is happy to announce the 2024 winner of the Gregor Reid Award for Outstanding Scholars in… -

Episode 27: Investigating the benefits of live dietary microbes
Investigating the benefits of live dietary microbes, with Prof. Colin Hill PhD and Prof. Dan Tancredi PhD Episode summary: In… -

Definition of postbiotics: A panel debate in Amsterdam
By Dr. Gabriel Vinderola, PhD, Associate Professor of Microbiology at the Faculty of Chemical Engineering from the National University of… -

Human milk oligosaccharides as prebiotics to be discussed in upcoming ISAPP webinar
Human milk oligosaccharides (HMOs), non-digestible carbohydrates found in breast milk, have beneficial effects on infant health by acting as substrates… -

ISAPP’s Guiding Principles for the Definitions of ‘Biotics’
By Mary Ellen Sanders, PhD, ISAPP Executive Science Officer Articulating a definition for a scientific concept is a significant challenge…. -

Do fermented foods contain probiotics?
By Prof. Maria Marco, PhD, Department of Food Science & Technology, University of California, Davis We frequently hear that “fermented… -
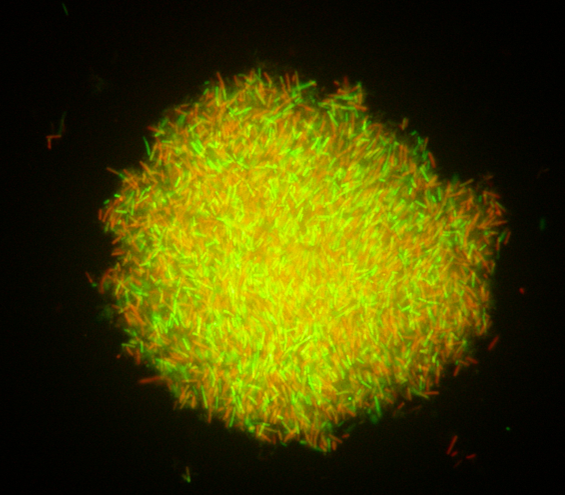
Should the concept of postbiotics make us see probiotics from a new perspective?
By Dr. Gabriel Vinderola, PhD, Associate Professor of Microbiology at the Faculty of Chemical Engineering from the National University of… -

Using probiotics to support digestive health for dogs
By Kelly S. Swanson, PhD, The Kraft Heinz Company Endowed Professor in Human Nutrition, University of Illinois at Urbana-Champaign, USA… -
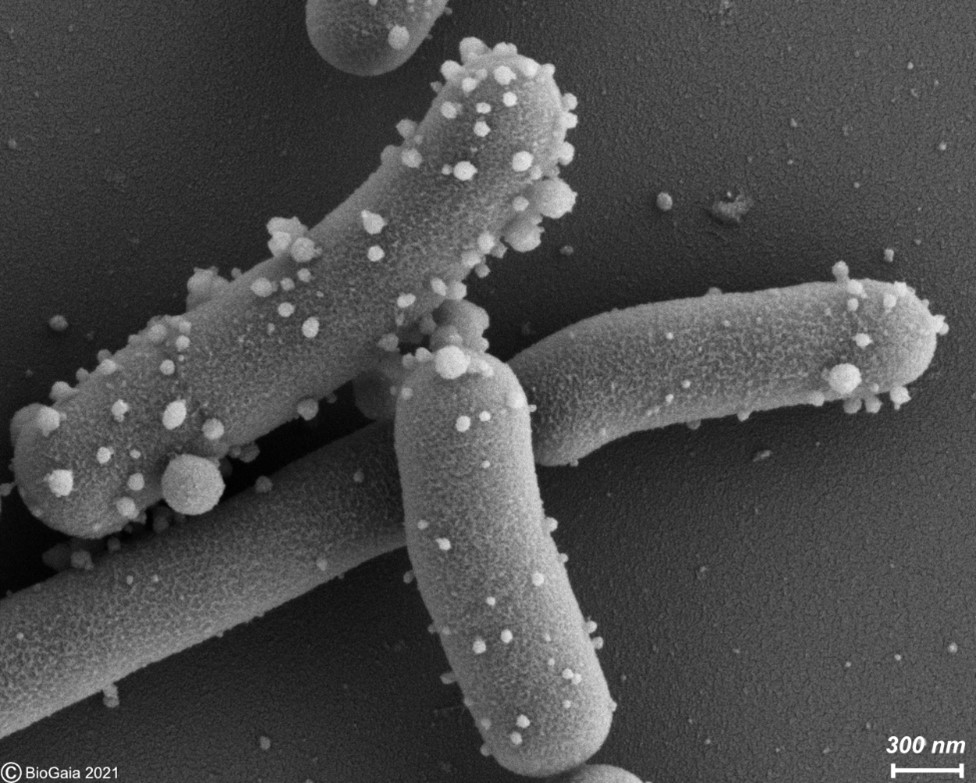
Bacterial vesicles: Emerging potential postbiotics
By Dr. Gabriel Vinderola, PhD, Associate Professor of Microbiology at the Faculty of Chemical Engineering from the National University of…
