1-2 of 2 results
-

Clarifying the role of metabolites in the postbiotic definition
By Dr. Gabriel Vinderola PhD, Instituto de Lactología Industrial (CONICET-UNL), Faculty of Chemical Engineering, National University of Litoral, Santa Fe,… -
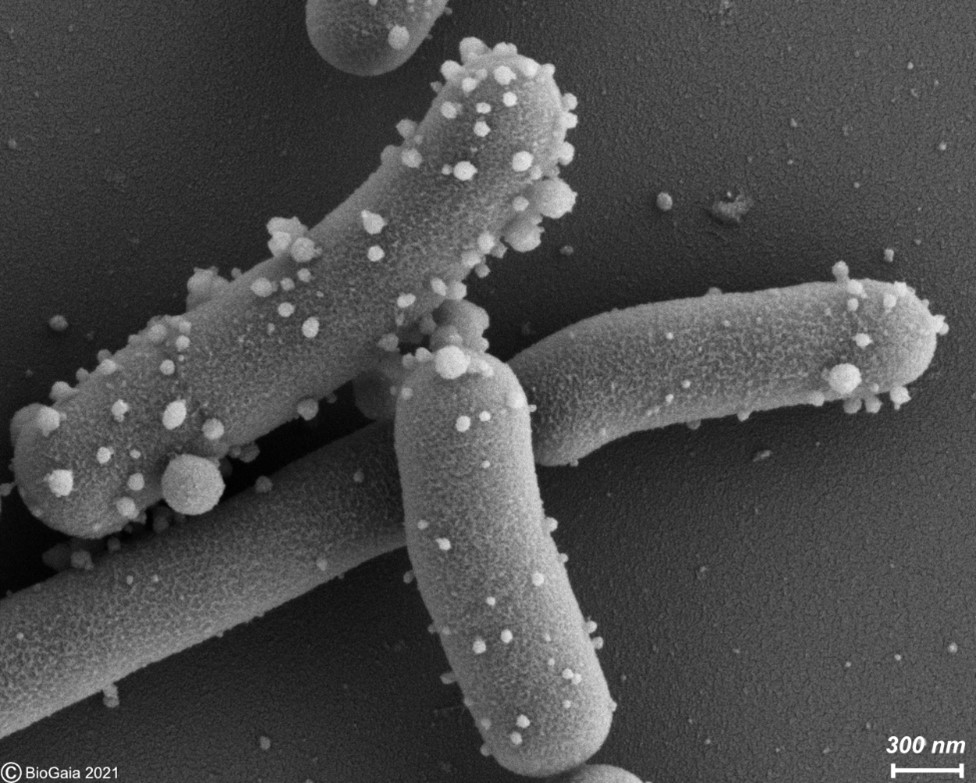
Bacterial vesicles: Emerging potential postbiotics
By Dr. Gabriel Vinderola, PhD, Associate Professor of Microbiology at the Faculty of Chemical Engineering from the National University of…